Uważa się, iż organelle takie jak mitochondria czy chloroplasty powstały na drodze endosymibiozy a ich prastrukturami były po prostu bakterie. Koncepcja endosymbiozy została po raz pierwszy zaproponowana przez Lynn Margulis w roku 1970. Swą teorie badaczka przedstawiła w książce po tytułem "The Origin ofthe Eucariotic CelI". Dziś teoria ta jest już uznanym biologicznym faktem i pokazuję jak ważnymi organizmami były w przeszłości bakterie.
Według teorii endosymbiozy autonomiczne organelle, jakimi są mitochondria oraz chloroplasty powstały z prokariontów, które w pewnym momencie stały się endosymbiontami innych organizmów. W przypadku mitochondrium punktem wyjściowym był tlenowy heterotrof, zaś chloroplastu tlenowy fototrof. To, co widzimy dziś kształtowało się na przestrzeni milionów lat, a zostało zapoczątkowane aktem "wchłonięcia" ówczesnej prymitywnej bakterii przez komórkę, u której zaczęła się tworzyć otoczka jądrowa. Bakteria ta nie uległa, jak to zazwyczaj bywa, procesowi trawienia lecz "zadomowiła" się na cytoplazmie stając się endosymbiontem komórki, która ją pochłonęła. Dzięki temu komórka ta zyskała bądź to zdolność do oddychania tlenowego (mitochondria), bądź do fotosyntezy (chloroplasty) albo do jednego i drugiego naraz, w zamian oferując ochronę oraz łatwy dostęp do substancji odżywczych.
Naukowcy szacują czas, w którym doszło do endosymbiozy na jakiś 1 mld lat temu. Od tamtego momentu obustronne korzyści bakterii i ich gospodarza przeistoczyły się w relację, która jest im niezbędna do życia. Doszło wręcz do tego, że do materiału genetycznego gospodarza dostały się fragmenty materiału bakterii kodujące ważne dla nich białka. Endosymbiotyczne struktury utraciły całkowicie zdolność do samodzielnego życia przekształcając się w organella.
Uważa się, że bakterie, które przekształciły się w mitochondria należały do rodzaju Paracoecus z grupy proteobacteria, zaś przodkami chloroplastów były sinice, które łączy z tymi strukturami zdolność do fotosyntezy.
Teoria Lynn Margulis uzyskała poparcie w badaniach genetycznych dotyczących materiału dziedzicznego organelli komórkowych oraz dzisiejszych przedstawicieli dzisiejszych prawdopodobnych grup wyjściowych dla tych struktur. Ponad to na poza komórkowe pochodzenie mitochondriów i plastydów świadczy wysoki stopień ich autonomii oraz to, że nowe struktury powstają nie "na nowo" lecz przez podział tych już istniejących. Posiadają one także własną maszynerię do produkcji własnych białek oraz (bardzo istotne) są zdolne do replikacji DNA się w nich znajdującego.
Na obszarze mitochondriów oraz plastydów występują cząsteczki DNA w postaci kolistej ( kolejna cecha bakteryjna) a ich rybosomy budową bardziej przypominają struktury znajdowane u bakterii, niż eukariotyczne rybosomy. Kolejne podobieństwo dotyczy błon, które otaczają organella i bakterie. Cechą wyróżniającą właśnie te struktury w komórce jest to, iż otacza je podwójna błona białkowo-lipidowa.
Lista takich dowodów jest dość długa i z biegiem czasów uzupełniana jest o kolejne punkty. Na koniec tego wyliczania może warto jeszcze tylko dodać, że mechanizmy odpowiedzialne za transfer genów od prokariotów do eucariotów są odnajdywane nawet jeszcze dziś u niektórych przedstawicieli eucariota. Przykładem mogą tu być chociażby dobrze nam znane drożdże, Sacharomyces cerevisiae.
Jak już wcześniej wspomniano niektóre komórki mogły wchłonąć tylko sinice lub tylko tlenowe heterotrofy. Analiza genomów znajdujących się w mitochondriach i plastydach potwierdziła przypuszczenia, iż "przodkowie" tych struktur wniknęli do komórki gospodarza w różnym czasie, a więc mamy tu do czynienia z dwoma, zupełnie niezależnymi aktami endosymbiozy.
W trakcie badań dotyczących powyżej omawianego tematu pojawiało się oczywiście wiele alternatywnych koncepcji. Na przykład według jednej z nich mitochondria a także chloroplasty powstały w wyniku wypuklenia się błon komórkowych do wnętrza komórki, które to po pewnym czasie utraciły ciągłość z reszta błon i stały się zupełnie autonomicznymi tworami.
Pomimo tego, że teoria endosymbiozy zdaję się być najlepszym wytłumaczeniem tego, co działo się miliard lat temu i ona nie pozbawiona jest wielu znaków zapytania. Pytania, które się przy jej okazji zrodziły dotyczą tych komórek pełniących funkcję gospodarza: jak radziły sobie te komórki zanim doszło do endosymbiozy? Poza tym jak dotychczas nie udało się odnaleźć, żadnych dowodów na istnienie takich tworów. Teorię te ciężko sprawdzi a jedyne co za nią przemawia to jej logiczność.
Klasyfikacja bakterii:
Długo zastanawiano się pod względem, jakiej cechy dałoby się stworzyć najlepszą i najwięcej mówiącą o bakteriach systematykę. W wszystkich opcji za najlepiej spełniającą ten wymóg, choć nie pozbawioną wad uznano klasyfikacje ze względu na aparat genetyczny, który posiadają bakterie.
Należy tu przypomnieć specyfikę tych organizmów, która to polega na tym, iż ich materiał genetyczny nie jest otoczony żadną błoną - brak zatem tzw. jądra komórkowego ( z łac. nucleus). Za to centralnie znajduję się spiralnie zwinięta nić DNA, której cząsteczka ma zazwyczaj postać kolistą. Strukturę tę określa się mianem nukleoidu.
Właśnie ze względu na te różnicę organizmy dzielimy na bezjądrzaste, które zwie się prokariotami (naukowo: Procaryota) oraz te, których materiał genetyczny osłania otoczka jądrowa złożona z dwóch błon i są to eukarionty ( naukowo: Eukaryota).
Podział nie kończy się jednak na tym, gdyż już w latach 70- tych Procaryota podzielono na dwie odrębne grupy archebakterie, inaczej Archaebacteria oraz bakterie właściwe, czyli Eubacterie. Obie te grupy łączy występowanie nukleoidu, lecz pod względem genetycznym bakteriom właściwym bliżej do eukariotów niż do archebakterii. Do takiego oto wniosku doszedł amerykański Carl Woese wraz ze swoim zespołem po przeanalizowaniu materiału genetycznego pochodzącego od "bakterii", które to znacznie różniły się od innych np. żyły w bardzo wysokich temperaturach, produkowały metan itp.
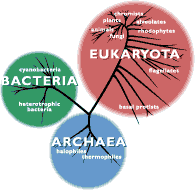
Systematyka świata żywego została przebudowana. W miejsce starej systematyki z 1959 roku autorstwa R. Whittakera według, której organizmy można było zaklasyfikować do jednego z pięciu królestw: Monera, Protista, Fungi, Plantae i Animalia wprowadzono nową. W jej ujęciu organizmy żywe można zaklasyfikować do jednej z trzech domen: Archae, Bacteria lub Eukaryota. Pozwała to na bardziej przejrzyste zobrazowanie ewolucji świata żywego, którą to zazwyczaj przedstawia się za pomocą tzw. drzewa filogenetycznego (rys.1). Z użycia wyszła też pierwotna nazwa archebakterii, która to sugerowała ich bliskie pokrewieństwo w grupą bakterii właściwych.
Środowisko występowania bakterii.
Właściwie to ciężko tu wymienić wszystkie środowiska, w których występują bakterie, gdyż te znajdują się praktycznie wszędzie! Ich liczbę szacuję się na jakieś 5 x 1030 ( liczbę taką zasugerował w swej pracy z 1998 roku profesor amerykańskiego pochodzenia Williama Whitman). Mimo tak szerokiego spektrum występowania warto chociażby trochę przedstawić warunki życia bakterii oraz ich do tych warunków przystosowania.
W stanie naturalnym nie znajdziemy miejsca, które nie byłoby zasiedlone przez jakąś grupę bakterii. Znajdujemy je w naszym jedzeniu, wodzie czy też powietrzu. Kiedy czytasz ten tekst miliony przedstawicieli tej "domeny" żerują w Twoim organizmie - jedne pomagają, drugie zaś są przyczyną wielu poważnych chorób. Znajdują się one m.in. w naszym przewodzie pokarmowy a także na naszej skórze. Jednak, do czasu, nie musimy się przejmować zbyt mocno ich obecnością, co więcej, aż 95% tych że bakterii nie stanowi dla nas większego zagrożenia a czasem są nam one wręcz niezbędne do zachowania dobrego zdrowia.
Inne bakterie zamiast ciał organizmów żywych zdecydowanie wolą środowisko nas otaczające. Często bywają one dość wybredne, więc nie wszystkie znajdzie się w jednym miejscu. Znamy bakterie, które bardzo dobrze czują się w glebie natomiast już w wodzie nie. Nie szkodzi to niczemu, gdyż nisza ta zostaje szybko wykorzystana przez innych krewniaków, którym środowisko wodne jak najbardziej odpowiada.
Ogólnie wyróżnia się 3 typy ekologiczne mikroorganizmów: wodne, glebowe oraz symbiotyczne. W tym ostatnim przypadku pojęcie "symbiotyczne" odnosi się do bardzo szerokiego pojęcia tego słowa.
1) Bakterie glebowe
Gleba jest środowiskiem bardzo złożonym, na które składają się takie czynniki jak związki mineralne oraz organiczne a także różne organizmy żywe, wszystko to nie jest rozmieszczone zbyt równomiernie, także występują obszary bogatsze i uboższe w pewne substancje. Taka skupiskowo - warstwowa budowa ma swoje odbicie oczywiście w rozmieszczeniu bakterii ją zamieszkujących. Wpływ na rozmieszczenie mają takie parametry jak wilgotność czy zasobność w składniki pokarmowe. Powodują one, iż liczebność bakterii w 1 gramie gleby może wahać się od kilku milionów do nawet miliarda! Aby bardziej uzmysłowić "wagę" tej sprawy wystarczy sobie wyobrazić, iż szacuje się, że całkowita masa bakterii żyjących w glebie wynosi około 100 trylionów ton! Gdyby rozłożyć je warstwą na powierzchni naszej planety osiągnęłaby ona grubość około 1,5 m! Według innych obliczeń w każdym akrze pola uprawnego ma znajdować się od 90 do 225 kg bakterii. Tą olbrzymią masę szczęśliwie dla nas stanowią przede wszystkim tzw. bakterie autochtoniczne tj. takie, które nie są dla nas groźne. Bakterie groźne jak chociażby laseczki tężca czy jadu kiełbasianego stanowią zaledwie ułamek promila wszystkich bakterii zamieszkujących glebę.
2) Bakterie zamieszkujące wodę
Środowisko wodne to potężny obszar. Wody na naszej planecie zajmują niemal 2/3 jej obszaru a liczbę bakterii je zamieszkująca szacuję się na około 25x 1029 jednostek. Podobnie jak w przypadku gleby toń wodna nie jest jednorodna. Parametrami pod kątem, których się różni są dostępność pokarmu, zasolenie, natlenienie czy tez temperatura. Badania wykazują, że w 1 ml wody, która zdaję się być czysta (np. pochodzącej z rzeki) można stwierdzić obecność prawie 1 mln komórek! Zaś w wodach zanieczyszczonych np. ściekach na 1 ml może przypadać nawet tysiąc raz więcej bakterii niż wyżej wymienione.
Jednym ze specyficznych środowisk związanym z wodą są muły denne. Stanowią one idealne siedlisko dla tzw. bakterii beztlenowych, ponieważ warunki tam panujące pozwalają im na przeżycie. Mikroorganizmy pojawiają się również w sąsiedztwie gorących źródeł oraz kominów hydrotermalnych znajdujących się na dnach oceanów, w okolicach których panują temperatury rzędu 270 do 380 °C. Zamieszkują również zbiorniki wodne o silnym zasoleniu (np. Morze Martwe), chociaż w tym wypadku liczebnością ustępują one raczej przedstawicielom Archae.
Kolejnym nietypowym miejscem są lodowce, które za swe siedlisko obierały bakterie psychrofilne, czyli te dla których optymalna temperatura do przeprowadzania procesów metabolicznych plasuję się niżej niż 15°C. Zazwyczaj optymalna temperatura ich wzrostu waha się w okolicach 0°C.
3) Bakterie symbiotyczne
Jeśli chodzi o ilość bakterii, które żyją w jakiś relacjach z innymi organizmami to liczbę tych, które zamieszkują układ pokarmowy wszystkich ludzi na świecie (6 miliardów) szacują się na około 4 x 1023 jednostek! Co więcej niemal 50% masy, która wypełnia naszą okrężnicę stanowią przede wszystkim pałeczki Escherichia coli! Także wnętrza organizmów innych zwierząt, zarówno tak swojskich jak krowy czy bardziej egzotycznych jak rurecznikowce (organizm zamieszkujący w pobliżu kominów hydrotermalnych) są miejscem życia dla różnych bakterii. Bydło posiada w swym układzie pokarmowym specjalną część żołądka przeznaczoną dla mikroorganizmów. Owa część nosi nazwę żwacza i wypełnia ją masa bakterii, które do życia nie potrzebują tlenu za to są zdolne do przetwarzanie celulozy. Przykładem tu mogą być takie bakterie jak Ruminococcus albus czy Fibrobacter succinogenes, dzięki których działalności celuloza zostaje rozłożona do cukru już przez bydło przyswajalnego jakim jest glukoza. W momencie, kiedy przeważającą komponentom pokarmu staje się skrobia zmianie ulega również skład gatunkowy bakterii znajdujących się w żołądku. Zaczynają dominować te osobniki, które dobrze radzą sobie z rozkładaniem skrobi np. Ruminobacter amylophilus. Gdyby nie te mikroorganizmy bydło nie było by wstanie przyswajać substancji pokarmowych pochodzenia roślinnego. Wytępienie antybiotykami tychże bakterii u krów, kóz czy też owiec wywołuję u nich serię poważnych schorzeń, co odbija się na ich produktywności.
Przetwarzanie materiałów pochodzenia organicznego
W każdej chwili na naszej ziemi są wytwarzane i deponowane ogromne pokłady materiałów pochodzenia organicznego tj. opadłe liście oraz obumarłe rośliny, szczątki zwierząt czy chociażby odpadki spożywcze pochodzące z ludzkich gospodarstw. Gdyby dużo wcześniej nie pojawiły się organizmy, które są wstanie przetworzyć a raczej rozłożyć i wprowadzić do ponownego obiegu elementów składowych tych substancji Ziemia z całą pewnością stała by się jednym wielkim śmietniskiem. Jak widać zatem zasług bakterii w tej kwestii nie da się przecenić!
Zacznijmy jednak po kolei. Za rozkład szczątków organicznych dostających się do gleby odpowiadają zamieszkujące ją mikroorganizmy. Nie są one na szczęście osamotnione w tym zadaniu, gdyż pomagają im tu również grzyby. Organizmy te wspólnie określa się mianem konsumentów jednak bardziej odpowiedni byłby tu termin "reducenci", który z resztą funkcjonuje w języku angielskim "decomposers". Dzięki działalności reducentów do ponownego obiegu dostaję się m.in. tak ważny pierwiastek jak węgiel, który wcześniej został uwięziony w bardziej złożonych związkach budujących ciała roślin czy też zwierząt. Dzięki mikroorganizmom zostaje on zredukowany do postaci, która może zostać łatwo przyswojona przez kolejne organizmy. Innym przykładem może być wędrówka azotu, pierwiastka bardzo ważnego dla całego świata żywego, który bez pomocy bakterii i grzybów tkwiłby uwięziony w pokładach gleby pod postacią np. nierozłożonych białek.
Bakterie są wstanie przetworzyć niemal każdy rodzaj związku organicznego, oczywiście nie wszystkie bakterie wszystkie związki ale poszczególni ich przedstawiciele poszczególne grupy związków. Tempo przetwarzania jest ściśle uzależnione od warunków panujących w środowisku oraz indywidualnych możliwości przerobowych jednostki. Znaczna frakcja materii organicznej jest rozkładana w wierzchnich pokładach gleby, gdzie panują bardzo dobre warunki tlenowe. Dlatego, aby gleba była dobrze użyźniona koniecznym jest jej stosunkowo częste spulchnianie.
Jeżeli chodzi o przerób i rozkładanie poszczególnych związków to pierwszą grupą, na której się skupimy będą białka. Związki te są złożone z wielu pierwiastków, poza tym mają zazwyczaj duże masy cząsteczkowe. Aby móc przystąpić do ich degradacji wpierw musza one zostać porozcinane na mniejsze fragmenty tj. polipeptydy, peptydy a potem aminokwasy. Odbywa się to poza komórkowo dzięki działaniu bakteryjnych lub grzybich proteaz.
Aminokwasy kolejno na drodze amonifikacji często przekształcane są do amoniaku, dzieję się tak gdyż uwalniane są z nich poszczególne grupy aminowe. Szkielet węglowy zaś bakterie wykorzystują do własnych przemian katabolicznych oraz anabolicznych. Zdarza się jednak, że owe grupy aminowe nie są przekształcane w amoniak ale na drodze transaminacji (przeniesienia) bywają wykorzystywane przez bakterie do ich własnych potrzeb.
Szczątki roślin obfitują w takie cukry jak celuloza czy lignina. W warunkach tlenowych celuloza rozkładana jest na związki prostsze prze bakterie tlenowe m.in. z rodzaju Cytophaga. Proces rozkładania takiego związku jak celuloza nie przebiega z byt szybko, ale jest on zdecydowanie szybszy niż rozkład ligniny, która zazwyczaj stanowi 30% suchej masy rośliny. Problem polega na złożoności budowy ligniny, którą to tworzą fenylopropany, czyli usieciowane związki zawierające w swej strukturze azot oraz pierścienie aromatyczne. Nie do końca wiadomo jak bakterie radzą sobie z rozkładem tego związku, wiadomo jednak że proces ten jest niezwykle ważny, gdyż stanowi podstawę tzw. kwasów humusowych.
W skład humusu wchodzi duża liczba trudno rozkładalnych związków, nieraz takich których nie da się zidentyfikować. Ponad to znajduję się on w stanie ciągłego "ruchu" - na jego obszarze zachodzą na przemian procesy tworzenia i rozkładu.
Jak już wspomniano wcześniej bakterie pełnia niezwykle istotną funkcję przy obrocie "węgla" w przyrodzie. Nie tylko rozkładają złożone związki organiczne do tych prostszych, ale także pozwalają wydostać się cząsteczką CO2 do atmosfery, gdzie mogą zostać przyswojone przez rośliny dzięki procesowi fotosyntezy. Chociaż z CO2 korzystać mogą również same bakterie tzw. autotroficzne, które wytwarzają niezbędną dla siebie energię w podobny sposób jak rośliny. Inną metodą może być chemosynteza, kiedy to energię pobiera się z przemian związków nieorganicznych.
Następnym pierwiastkiem, którego nieustanna cyrkulacja jest bardzo istotna dla świata żywego jest azot. Dzięki unikalnym zdolnościom mikroorganizmów do gleby dostaję się on w postaci łatwo przyswajalnej dla roślin, bądź też uwalniany jest do atmosfery w postaci nieczynnej chemicznie N2. Jednak to jak ponownie odzyskać azot z powietrza prześledzimy poniżej.
Jak bakterie wiążą azot atmosferyczny?
Zanim zaczniemy omawiać ten problem należy zaznaczyć, iż bakterie wraz z niektórymi archeonami są jednymi organizmami, które są wstanie przyswajać oraz wbudowywać w swe struktury azot w postaci cząsteczkowej. Wydaję się to być strasznym marnotrawstwem z punktu widzenia roślin, gdyż powietrze stanowi ogromny magazyn tegoż pierwiastka ( ok. 70%) jakże dla nich niezbędnego. Rośliny musiały zatem wejść w odpowiednie relacje z bakteriami, które to posiadają zdolność przyswajania tej postaci azotu, by móc wyrównywać sobie na tej drodze jego niedobór. Bakterie o takich zdolnościach stały się niezwykle ważną grupą z punktu widzenia obiegu tego pierwiastka w przyrodzie a zamieszkują one przede wszystkim gleby, co jest jak najbardziej zrozumiałe.
Dokonuję się pewnego podziału tejże grupy bakterii na te, które występują w stanie niezwiązanym - należą do nich organizmy z rodzajów Azotobacter czy Clostridium, oraz na te, które żyją jedynie dzięki symbiozie z roślinami - przedstawicielem tu mogą być chociażby osobniki z rodzaju Rhizobium.
Mimo faktu, że wiązanie azotu atmosferycznego jest tak niezwykle ważnym procesem równocześnie wiąże się on z wysokimi kosztami energetycznymi. Już na wstępie trzeba "użyć" jej niezwykle dużo( na 1 cząst. N2 potrzeba aż 16 cząst. ATP), aby rozerwać potrójne wiązanie między dwoma atomami azotu. Energia ta zazwyczaj pochodzi z różnych procesów metaboliczny w zależności od mikroorganizmu. Te, które same nie są wstanie wytworzyć produktów do wysokoenergetycznego oddychania pobierają je od swoich partnerów tj. roślin.
Na poziomie molekularnym bakterie swą zdolność do wiązania N2 zawdzięczają specyficznemu enzymowi o nazwie nitrogenaza, która to w swoim składzie posiada między innymi pierwiastek molibdenu oraz cechują ją niezwykłą wrażliwość na tlen. Dlatego tez wiązanie azotu może zachodzić jedynie w warunkach beztlenowych! Białkiem towarzyszącym jest tzw. reduktaza nitrogenazy. Jeżeli chcielibyśmy założyć hodowlę takich bakterii najwdzięczniejszym obiektem jest tu Azotobaeter, który jest bakterią niesymbiotyczną, czyli nie potrzebującą partnera. Wystarczy pobrać nieco gleby i położyć je na odpowiednim podłożu mikrobiologicznym, które pozbawione jest azotu, ale za to bogate w źródło węgla a następnie przez pewien czas inkubować.
Jeśli zaś chodzi o te bakterie, które wiążą azot, ale potrzebują one do istnienia partnera to znajdują się one zazwyczaj w brodawkach korzeniowych - specjalnych strukturach wytwarzanych przez rośliny w celu stworzenia odpowiedniego środowiska bytowania dla bakterii. Najbardziej znanym przykładem symbiozy bakterii przyswajających azot i roślin są gatunki roślin motylkowych tj. łubin czy groch. W wytworzonych na ich korzeniach brodawkach bakterie ulegają przekształceniu w tzw. bakterioidy.
Relację tego typu są niezwykle ważne z punktu widzenia m.in. rolnictwa. Przy odpowiedniej znajomość biologii roślin i bakterii rolnicy są wstanie uzyskiwać wysokie plony nawet na glebach ubogich w azot. Co więcej rośliny motylkowe stają się po okresie wegetacyjnym doskonałym nawozem wzbogacającym tą że glebę od deficytowy pierwiastek. Sytuacja ta jest wykorzystywana głównie przez rolników, którzy prowadzą wysiewy roślin motylkowych w postaci poplonów, które później są zaorywane.
Mimo niezwykłych korzyści wynikających z wiązania wolnego azotu sposób ten jest niezbyt wydajny i dlatego rzadko stosowany, gdy są inne możliwości dostarczenia roślinom azotu.
Proces nitryfikacji
Do zupełnie innego rodzaju procesów dotyczących przemian azotu w przyrodzie zaliczamy nitryfikacje. Jego istota opiera się na przemianie amoniaku bądź jego soli na drodze utleniania do azotanów. Podobnie jak wcześniej omawiane grupy ekologiczne bakterii tak i te głównie zasiedlają glebę. Energię potrzebną im do asymilacji CO2 pozyskują dzięki dwuetapowemu już wcześniej wspomnianemu utlenianiu amoniaku (bądź soli amonowych) wpierw do azotynów, a później do azotanów. Każdy z tych etapów przeprowadzany jest przez zupełnie inne bakterie i tak na pierwszym etapie, kiedy to NH4+ jest przekształcane do NO2- działają głównie bakterie z rodzaju Nitrosomonas, zaś później od NO2- do NO3- Nitrobacter. Związane są one ze sobą silną współpracą, gdyż azotowy produkt pierwszych jest silnie toksyczny i musi ulec błyskawicznej przemianie do postaci nieszkodliwej.
Bakterie przeprowadzające nitryfikację są tlenowcami, toteż w wyniku ich procesów metabolicznych powstają cząsteczki CO2 a azot zostaje przekształcony do postaci niezbyt korzystnej dla roślin. Kolejnym minusem jest owo powstawanie azotynów, które jeśli nie zostaną szybko wykorzystane są wypłukiwane przez wodę i tracone.
Obieg siarki
Pierwiastek ten obok węgla i azotu stanowi bardzo ważny element biogenny. Przez większość bakterii i roślin jest on przyswajany w postaci jonowej ( SO42-), chociaż nie jest to jedyna postać tego pierwiastka, która może być wykorzystywana. Nieocenioną rolę pełnią np. mikroorganizmy, które są wstanie utlenić siarkę nawet w jej najmocniej zredukowanej postaci siarkowodoru (związek niezwykle toksycznego). Jak na razie znane są nauce dwie grupy bakterii, które potrafią utleniać związki siarki. Są to bakterie siarkowe, które dzięki utlenianiu siarki uzyskują energię do asymilacji CO2- przykładem może tu być Thiobacil hiooxidans oraz bakterie fotosyntetyzujące, które nie potrzebują tlenu a siarka stanowi źródło elektronów (jest ich donorem). Bakterie te spotyka się głównie w wodzie, chociaż można się na nie natknąć i w glebie.
Neutralizacja związków toksycznych
Wszystkie opisane procesy metaboliczne należą do codziennych "obowiązków" bakterii. Jednak zdarza się i tak, że w środowisku pojawiają się jakieś związki "nietypowe", które mogą stanowić duże dla niego zagrożenie. Wtedy to bakterie okazują się równie przydatne jak na co dzień.
Na przykład w wyniku jakieś awarii bądź po prostu ludzkiej głupoty do środowiska dostają się takie toksyny jak ropa, benzyna czy jakieś związki policykliczne. Dzięki występowaniu w glebie mikroorganizmów, które są wstanie zdegradować takie substancję nie każda taka sytuacja prowadzi do tragedii ekologicznej.
Korzystny wpływ na strukturę ziemi mają np. bakterie z grupy promieniowców (Actinomycetes) to właśnie ich obecności zawdzięczamy ten unikatowy zapach, jaki posiada nasza ziemia.
Praca na bazie pozycji: "Świat bakterii" autorstwa Mirosława Włodarczyka
Rys. 1 źródło
http://www.ucmp.berkeley.edu/archaea/archaea.html
