Gen
Tym mianem określamy odcinek cząsteczki kwasu deoksyrybonukleinowego (DNA) kodujący informację na temat budowy jednej cząsteczki peptydu lub jednego łańcucha białka. Geny ułożone są liniowo w chromosomach (w przypadku bakterii: w chromosomie). Rodzaj i dobór genów są specyficzne dla gatunku. Mutacje, czyli zmiany chemiczne w strukturze cząsteczek DNA, dostarczają nowych, "alternatywnych" odmian genu, czyli alleli genu. Organizmy o podwójnym garniturze chromosomowym, takie jak człowiek, większość zwierząt i roślin mają po dwa allele dla każdego genu te same lub różne. Tak więc ta sama cecha może być kodowana przez różniące się od siebie geny, czego przykładem są owłosione lub gładkie strąki grochu. O tych dwóch genach powiemy, że są to geny alleliczne (alternatywne, wykluczające się). Geny owłosienia i np. długości strąku określimy jako niealleliczne, gdyż strąk może być równocześnie długi i owłosiony (lub nieowłosiony).
Informacja genetyczna
Informacja dziedziczna zapisana jest w DNA za pomocą kodu genetycznego, określając strukturę białek, a wraz z nią wszystkie uchwytne w obserwacji cechy organizmu. Ludzie od dawna zastanawiali się nad tym, w jaki sposób z malutkiego zarodka rozwija się dorosły organizm. W czasach istniały obozy zwolenników preformacji i epigenezy. Obóz preformacji głosił, że kompletny organizm jest zawarty w plemniku lub jaju jako miniaturka dorosłego. W komórkach, z których rozwiną się gamety, zawarty jest "wzorzec" potomstwa, jak mniejsza ruska wańka-wstańka w większej. Głosiciele teorii epigenezy forsowali pogląd, że zarodek powstaje jednorazowo i całkowicie od nowa. Podobieństwo potomka do obojga z rodziców tłumaczono "mieszaniem się ich krwi" w równych proporcjach. Od dawna już dostrzegano podobieństwo, czy nawet tożsamość zjawisk dziedzicznych zachodzących u człowieka i u reszty świata żywego. Dzisiejsza wiedza po części przyznaje rację każdemu z tych dawnych obozów. Nikt w XVIII wieku by nie przypuszczał, że głęboko we wnętrzu komórki ukryta jest niewidoczna nitka kwasu DNA i istotnie młody, rozwijający się organizm realizuje ukryty w niej, przekazywany od pokoleń schemat. Jednocześnie z jednej komórki powstają biliardy całkiem nowych komórek, które różnicują się na wiele typów, i organizują w tkanki i narządy. Informacja genetyczna istotnie "przelewa się" z rodziców na potomków, i istotnie posiadamy po połowie genów od ojca i od matki, choć "zlewanie się" obejmuje molekularne procesy, o których jeszcze w XIX wieku nikt nie miał pojęcia, crossing-over, losowe rozchodzenie się chromosomów i powstawanie jądra ze zlanych przedjądrzy. Trafnie dostrzeżono podobieństwo tych procesów w zwierząt i roślin, choć u tych ostatnich embriogeneza zatrzymuje się na jakiś czas, aby zarodek mógł przebyć stadium spoczynkowe, konieczne do przeczekania trudnych warunków atmosferycznych i braku światła, efektywniej rozprzestrzenić się w okolicy. Blokada przestaje działać pod wpływem wilgoci, temperatury, światła.
Przyrodników nurtowało pytanie, jakie to materialne czynniki dziedziczności są "przelewane" z rodziców na dzieci, przenosząc pierwiastki wzajemnego podobieństwa. Na pytanie to po części odpowiedział Grzegorz Mendel, zakonnik żyjący i pracujący w klasztorze augustianów w Brnie na Morawach. Początkowo jako osoba odpowiedzialna za ogród klasztorny prowadził doświadczenia z hodowlą czystych linii grochu, które podsunęły mu pomysł późniejszych doświadczeń. Mianem linii czystych określamy zbiory okazów hodowanych i krzyżowanych między sobą tak, że osobniki utrzymują się w typie (np. pod względem barwy kwiatów). Mendel hodował wiele linii czystych, np. pod względem nasion gładkich lub pomarszczonych, zielonych bądź żółtych, o łodygach wysokich lub niskich, strąkach owłosionych lub nagich. Zaobserwował, że osobniki "czyste" pod względem jednej cechy nie muszą, choć mogą być czyste również pod względem innych. Kolejnym ważnym krokiem stało się skrzyżowanie dwóch osobników, pochodzących z alternatywnych linii czystych. Okazało się, że potomstwo przejawiło cechę tylko jednego z rodziców. W następnym pokoleniu jednakże rozkład cechy wynosił już 3 (jednego z rodziców) :1 (cecha drugiego z rodziców). Cechę obecną w pierwszym pokoleniu mieszańców i przeważającą w drugim Mendel nazwał dominującą, a cechę alternatywną - recesywną. Zakonnik prowadził swoje badania nadzwyczaj skrupulatnie, potwierdzając je ogromną liczbie krzyżówek i ściśle trzymając się matematycznych obliczeń. Będąc pewnym prawdziwości swych odkryć, po latach i setkach udanych doświadczeń zdecydował się publicznie przedstawić swoje wyniki. Jego rewelacje trafiły na moment, gdy cały świat przyrodniczy fascynował się raczej mikroświatem ujawnianym przez coraz doskonalsze mikroskopy, nowymi mikrobami, odkryciami nieznanych gatunków zwierząt i zgłębianiem fizjologii i anatomii człowieka. Obserwacje poczynione na hodowli grochu nie wzbudziły niczyjego zainteresowania. Dopiero burza rozpętana przez teorię Darwina i Wallace'a oraz odkrycie chromosomów i ich roli w dziedziczności sprawiło, że prace Mendla ujrzały światło dzienne, a jego prawa istotnie uzupełniły kształtujący się obraz świata istot żywych.
Genialne odkrycie Mendla dotyczyło obecności dwóch zawiązków istniejących dla każdej cechy w komórce. Zwykle cesze nadaje kształt jeden z zawiązków. Trafiają one do gamet pojedynczo, niezależnie od siebie podczas oo- i spermatogenezy. Łączące się w zygotę plemnik i komórka jajowa restytuują podwójną liczbę zawiązków cechy. Zawiązki są niezmiennymi całościami, nie mieszającymi się, zachowującymi jak dwa oddzielne byty, nie tracące swojej tożsamości w momencie wchodzenia do gamet ani też ich łączenia się znowu w funkcjonujący organizm. Ponieważ organizm produkuje tysiące (żeński) lub miliardy gamet (męski), mechanizmy dbające o dokładność powielania zawiązków cech muszą być nadzwyczaj sprawne. Pod koniec XIX w. odkryto organelle, których zachowanie w trakcie podziału komórkowego odpowiadało postulowanemu zachowaniu się "zawiązków cech". Były nimi chromosomy. Są to małe, pałeczkowate struktury występujące w jądrze komórkowym, uwidaczniające się w trakcie jego podziału na dwa potomne (mitozy lub mejozy). Podział komórki najbardziej elementarnym procesem rozmnażania. Potomstwo podobne jest do komórki macierzystej jak dwie krople wody. Każda komórka potomna otrzymuje bowiem "w spadku" po jednym pełnym komplecie i chromosomów, i zawiązków wszystkich cech. Zachowanie się chromosomów w trakcie tworzenia gamet było całkowicie zgodne z postulatami Mendla. Gameta otrzymuje po połowie garnituru chromosomów, natomiast zygota odtwarza liczbą podwójną.
Dzisiejsza wiedza o chromosomach jest większa i dokładniejsza, niż przed laty. Zbudowane są z dwóch klas związków organicznych, kwasów nukleinowych i białek. W 1944 r. amerykański biochemik Oswald Avery ostatecznie udowodnił, że to kwas deoksyrybonukleinowy jest materialnym nośnikiem informacji dziedzicznej. Nieprecyzyjne pojęcie "zawiązków cech" stawało się coraz bliższe wiedzy empirycznej. Stwierdzono, że wystawiając komórkę na działanie promieniowania UV, ciepła czy pewnych substancji można wywołać zmiany jej właściwości, a także wszystkich komórek od niej się wywodzących. Okazało się, że informacja o budowie organizmów ma podobny związek ze swym materialnym nośnikiem, jak poemat lub przepis kulinarny z zadrukowaną kartką książki. Poemat nie ma jednak "zastosowania", natomiast jedynym celem zapisu informacji genetycznej jest sterowanie rozwojem i procesami życiowymi organizmu.
Odkrycia Mendla z perspektywy XXI wieku.
Przyczyn rozmaitości cech u potomstwa pozornie jednolitych rodziców dociekano na długo przed Mendlem. Jedno ze starszych źródeł pisanych świata, Księga Rodzaju, zawiera z naszego punktu widzenia bardzo naiwną analizę tych zjawisk: "Jakub nazbierał sobie świeżych gałązek topoli, drzewa migdałowego i platanu i pozdzierał z nich korę w taki sposób, że ukazały się na nich białe prążki. Tak ostrugane patyki umocował przy korytach z wodą, czyli przy poidłach, aby je widziały trzody, które przychodziły pić wodę. Gdy bowiem zwierzęta przychodziły pić wodę, parzyły się. I tak parzyły się zwierzęta z trzód przed tymi patykami i wskutek tego dawały przychówek o sierści prążkowanej, pstrej i cętkowanej".
O ile dalsze wieki rozwoju nauki przyniosły lepsze rozumienie dziedziczenia, Mendel dostarczyły trwałej podstaw teoretycznych dla tych zjawisk. Zawdzięczamy mu uzyskanie i pielęgnację linii czystych oraz dokładne przeliczanie wyników i zaopatrzenie ich w uwagi natury statystycznej. Poniższa tabela zawiera przykład danych, jakimi posługiwał się Mendel:
|
krzyżówka rodzicielska
|
fenotyp F1
|
stosunek fenotypów w pokoleniu F2
|
stosunek F2
|
|
nasiono gładkie x pomarszczone
|
gładkie
|
5474 gładkich:1850 pomarszczonych
|
2.96:1
|
|
nasiona żółte x zielone
|
żółte
|
6022 żółtych:2001 zielonych
|
3.01:1
|
|
kwiaty czerwone x białe
|
czerwone
|
705 czerwonych:224 białych
|
3.15:1
|
|
wysokie x niskie rośliny
|
wysokie
|
787 wysokich:227 niskich
|
2.84:1
|
Tabela 1. Wyniki liczbowe krzyżówek grochu pachnącego.
Kilka terminów użytych w tej tabelce może być niezrozumiałych. Pod pojęciem fenotypu, (dosłownie 'to, co jest możliwe do obserwacji') rozumiemy zewnętrzny, fizyczny wygląd danej cechy. W tabelce powyżej mamy fenotypy kwiatów białych i kwiatów czerwonych, nasion żółtych i pomarszczonych oraz roślin wysokich i niskich. Pod pojęciem pokolenia F1, F2 itd. rozumiemy kolejne pokolenia powstałe z krzyżowania osobników grochu, których nazwa bierze się z łacińskiego filius, syn. Przeanalizujmy teraz dane zawarte w tabelce, choć to będzie trochę kłopotliwe. Wysiłek jednak opłaci się, bowiem pozwoli nam zrozumieć zjawiska dziedziczne bardzo ściśle i jednoznacznie ujęte.
W pokoleniu F1 pojawia się, jak widzimy w drugiej kolumnie tabeli, tylko jeden z dwóch fenotypów rodzicielskich. Rołsiny F1 "noszą w sobie" informację genetyczną, która już w kolejnym pokoleniu da nam wszystkie znane u rodziców fenotypy. Pokolenie F2 wykazuje stosunek fenotypów 3:1, przy czym cecha dominująca pojawia się trzy razy tak często, jak recesywna. Monohybrydą nazwiemy potomka dwojga rodziców, którzy są homozygotami alternatywnych alleli pary genowej. Monohybrydy są dogodne do opisu relacji pomiędzy allelami. Mendel ukuł dwa terminy w celu opisania relacji dwóch alleli, oparłszy je na obserwacjach fenotypów uzyskiwanych w pokoleniach F1 i F2. Jako allel dominujący określił ten allel, który ulega ekspresji kosztem allelu alternatywnego; określa on fenotyp, który jest widoczny w pokoleniu F1 powstałym ze skrzyżowania różnych linii czystych. Dominacją nazywać będziemy zdolność jednego allelu do ekspresji fenotypu kosztem allelu alternatywnego; jest to najczęstsza forma interakcji pomiędzy allelami w przyrodzie. Zazwyczaj allel dominujący odpowiada za produkt, którego recesywny nie potrafi stworzyć, ulega zatem ekspresji, gdziekolwiek się pojawia. Allelem recesywnym jest ten, którego ekspresja jest tłumiona pod obecność allelu dominującego; określa fenotyp, który znika w pokoleniu F1 wynikłym z krzyżowania dwóch czystych linii, a pojawia się znowu w pokoleniu F2. Gdy allel jest homozygotyczny, powinien wykazać swój fenotyp. Fenotyp heterozygoty pozwala więc na określenie wzajemnej relacji alleli.
Ważne konsekwencje praw Mendla
Podsumujmy, co wiemy z dotychczasowych przemyśleń:
1. Czynniki dziedziczne mają charakter cząsteczkowy. Cząsteczki te nazywamy genami.
2. Każde z rodziców wnosi jedną parę genu do każdej komórki nowego organizmu, dla każdej z badanych cech.
3. F1 ze skrzyżowania dwóch linii czystych zawiera po jednym allelu warunkującym fenotyp dominujący i recesywny, tworzące parę genów.
4. Zawsze jeden allel z pary genów segreguje do każdej z tworzonych gamet, stąd też gameta przenosi tylko jeden allel z każdej pary.
5. Gamety łączą się losowo, a geny dobierają w pary niezależnie jeden od drugiego.
Definicje w genetyce Mendlowskiej.
Utrwalmy teraz definicje genetyki Mendlowskiej.
Allel - alternatywna forma danej pary allelicznej; dla każdego konkretnego genu mogą istnieć więcej niż dwa allele, lecz w jakimkolwiek osobniku mogą znajdować się równocześnie dwa allele (para genów).
Para alleliczna - kombinacja dwu alleli dla tego samego genu, jakie posiada osobnik.
Homozygota - osobnik, który posiada tylko jeden allel w parze allelicznej. Przykładowo DD jest homozygotą dominującą, posiadającą tylko jeden allel D; dd jest homozygotą recesywną z allele d.
Heterozygota - osobnik, który posiada dwa allele tej samej pary genów; przykładowo Dd.
Genotyp - specyficzna kombinacja alleli.
Więcej genetyki Mendlowskiej.
Używając odpowiednich symboli możemy zilustrować krzyżówkę wysokich i niskich roślin grochu w następujący sposób:

Pokolenie F2 uzyska się przez samozapylenie roślin F1. Można to graficznie zilustrować w postaci kwadratu Punnetta. Bazując na tego typu wynikach Mendel ukuł szereg innych terminów i sformułował swe pierwsze prawo. Na początku pokażę jednak kwadrat Punnetta:
|
D
|
d
| |||
|
D
|
DD
(wysokie)
|
Dd
(wysokie)
| ||
|
d
|
Dd
(wysokie)
|
Dd
(niskie)
|
Kwadrat Punnetta pozwala na określenie specyficznych stosunków genetycznych:
(1) rozszczepienie genotypów w F2: 1 DD : 2 Dd : 1 dd (inaczej 3 D_ : 1 dd)
(2) rozszczepienie fenotypów w F2: 3 wysokie : 1 niskie
Pierwsze prawo Mendla zwane jest prawem segregacji; podczas formacji gamet każdy członek pary alleli oddziela się od drugiego, a następnie współtworzy genetyczny obraz gamety. Prowadząc swe obserwacje, Mendel mógł sformułować hipotezę o segregacji. By ją sprawdzić, krzyżował ze sobą rośliny pokolenia F2. Jeżeli prawo było prawdziwe, mógł próbować przewidzieć skutek takiej krzyżówki. I faktycznie, wyniki były się takie, jak tego oczekiwał Mendel:
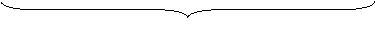 |

Na podstawie tych wyników znalazł potwierdzenie genotypów osobników pokolenia F2. Stosunek wynosi 1/4 DD + ½ Dd + 1/4 dd = 1.
Mendel wykonał jeszcze inną krzyżówkę, by potwierdzić swą hipotezę o segregacji. Była to krzyżówka wsteczna. Pamiętamy, że pierwsza krzyżówka dotyczyła rodziców pochodzących z linii czystych i dawała w efekcie heterozygoty F1:

W tym momencie zamiast samozapylenia F1, Mendel skrzyżował F1 z czystą linią homozygotycznych roślin niskich:
Krzyżówka wsteczna: Dd x dd
|
Gamety męskie
| ||
|
d
| ||
|
Gamety żeńskie
|
D
|
DD
(wysokie)
|
|
d
|
dd
(niskie)
|
Podsumowując, krzyżówka wsteczna przyniosła fenotypy w proporcji 1 wysokich : 1 niskich.
Krzyżówką wsteczną nazwiemy krzyżówkę hybrydy F1 z jednym z homozygotycznych rodziców; w przypadku wysokiego grochu krzyżówka może mieć postać Dd x DD lub Dd x dd; bardzo często jednak w krzyżówce wstecznej udział bierze osobnik rodzicielski recesywny. Krzyżówka testowa jest to krzyżówka jakiegokolwiek osobnika z osobnikiem rodzicielskim o cechach recesywnych; używa się jej do określenia, czy osobnik jest homozygotą dominującą, czy heterozygotą. Jak dotąd, nasza dyskusja koncentrowała się na krzyżówkach monohybrydowych. Nazywamy tak krzyżówkę między rodzicami, których różni od siebie pojedyncza para genów (zwykle AA x aa).
Genetyka w zadaniach i przykładach.
Zadanie 1. Skrzyżowano mysz długouchą z myszą krótkouchą.
F1: wszystkie myszy długouche.
F1 x F1 → F2: 36 długouchych i 13 krótkouchych (~ 3:1).
Uwaga: zapis 3:1 dotyczy frekwencji, czyli prawdopodobieństwa pojawienia się u potomstwa danej cechy. Prawa Mendla nie przestałyby funkcjonować, gdybyśmy otrzymali np. 24 myszy długouche i 20 krótkouchych, jednak najczęściej liczebności mają się do siebie mniej więcej jak 3:1.
Wniosek: allelem dominującym jest allel długich uszu u myszy.
Zadanie 2. Małżeństwo osoby z grupą krwi A z osobą z grupą krwi B.
F1: wszystkie dzieci grupa AB.
Jedno z dzieci znajduje partnera również z grupą krwi AB.
F1 (AB) x F1 (AB) → F2
F2: grupa A = 2 dzieci, grupa AB = 4 dzieci, grupa B = 1 dziecko (~ 1:2:1).
Wniosek: typy A i B warunkują osobne allele. Brak dominacji, allele kodominują (AB).
Zadanie 3. Krzyżujemy myszkę bezogonową z myszką normalną.
F1: 10 bez ogona, 9 normalnych (~ 1:1)
F1 (bezogonowa) x F1 (bezogonowa) → F2
F2: 10 normalnych, 21 bezogonowych, 9 martwych (~ 1:2:1)
Wniosek: gen warunkujący brak ogona w formie homozygotycznej jest letalny, to znaczy śmiertelny dla swoich nosicieli.
Zadanie 4.
Diagram obrazuje pokrewieństwa:
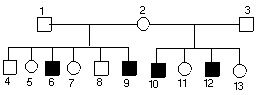
Mężczyźni oznaczeni są kwadratami, kobiety kółkami, osoby zdrowe na biało, obciążone chorobą genetyczną na czarno. Widać, że chorują tylko mężczyźni, zatem choroba musi być dziedziczona w chromosomie X.
Zadanie 5. Krzyżujemy czerwonookie muszki owocowe (♀) z muszkami czerwonookimi (♂)
F1: 50 czerwonookich (♀), 25 czerwonookich (♂), 25 białookich (♂)
Krzyżujemy F1 (białooki ♂) x F1 (czerwonooka ♀) → F2
52 krzyżówki F2 dały: 30 czerwonookich (♂), 33 czerwonookie (♀)
48 krzyżówek F2 dało: 22 czerwonookie (♂), 24 czerwonookie (♀), 21 białookich (♂), 23 białookie (♀).
Wniosek: gen barwy oka zlokalizowany jest u muszek na chromosomie płciowym X, dlatego ♂ nigdy nie przekazuje genu barwy oka potomstwu ♂.
Zadanie 6: szlaki metaboliczne. Załóżmy istnienie szlaku metabolicznego prowadzącego do produkcji barwnika u roślin:
enz A enz B
składnik 1 ----------> składnik 2 ------------> składnik 3
(bezbarwny) (różowy) (purpurowy)
Załóżmy też, że nosiciele alleli A i B produkują funkcjonalny enzym, podczas gdy nosiciele a i b nie produkują go. Przyjmijmy, że jedna funkcjonująca kopia enzymu wystarcza do katalizy reakcji.
Krzyżując AaBb X AaBb, ustal kolory potomstwa.
Odpowiedź: możliwe genotypy i fenotypy u potomstwa to 1 x AABB (purpurowy), 2 x AABb (purpurowy), 1 x AAbb (różowy), 2 x AaBB (purpurowy), 4 x AaBb (purpurowy), 2 x Aabb (różowy), 1 x aaBB (bezbarwny), 2 x aaBb (bezbarwny), 1 x aabb (bezbarwny) èpurpurowy : różowy : bezbarwny = 9 : 3 : 4
Załóżmy teraz, że szlak metaboliczny jest zorientowany alternatywnie:
enz A enz B
czerwony <----------bezbarwny składnik--------------> niebieski
barwnik barwnik
i wykonajmy taką krzyżówkę:
Aabb X aaBb
F1: 4 x AaBb (letalny), 4 x Aabb (czerwony), 4 x aaBb (niebieski), 4 x aabb (biały) è
czerwony : niebieski : biały = 1 : 1 : 1
Budowa i właściwości DNA
Swoje zastosowanie w kodowaniu informacji zawdzięcza DNA stosunkowo prostej budowie chemicznej. Cząsteczka DNA jest łańcuchem (nicią), w każdej komórce człowieka jest łącznie ok. 1 metra DNA. "Łańcuch" zbudowany jest z "ogniw". Ogniwa są nukleotydami, to jest kompleksami reszty fosforanowej H2PO4-, pięciowęglowego cukru (deoksyrybozy) i jedno- lub dwupierścieniowej zasady azotowej, tyminy (T), cytozyny (C), adeniny (A) bądź guaniny (G). Rodzaj podstawionej do nukleotydu zasady azotowej okazał się literą alfabetu, w jakim zapisana jest informacja genetycznej. Niezliczone możliwości kombinacji nukleotydów wzdłuż nici kodują informacje o wyglądzie i właściwościach tak skomplikowanego tworu, jak organizm człowieka ze wszystkimi jego narządami, w tym z mózgiem, sposobach zachowań i reakcjach na bodźce. Komórka posiada molekularną maszynerię, budującą na podstawie DNA konstrukcje z białka. W 1953 r. James Watson i Francis Crick, chemicy pracujący wspólnie w laboratorium w Cambridge w Anglii stali się odkrywcami trójwymiarowej struktury DNA. Nić kodująca informację sprzężona jest z drugą nicią. Wspólnie skręcają się w spiralę, fachowo nazywaną podwójną helisą (dubble helix). Obie nici utrzymywane są wzajemnie razem dzięki energii wiązań wodorowych pomiędzy zasadami azotowymi. Adenina potrafi w tej konfiguracji, w jakiej włączona jest do DNA, parować się tylko z tyminą, a cytozyna z guaniną. Kolejność zasad azotowych w jednej nici jest w pewnym sensie "odbiciem lustrzanym" drugiej, i tylko jedna może przechowywać sensowną informację o białkach (zwana nicią kodującą), podczas gdy druga tylko stabilizuje pierwszą (nić niekodująca). Materiał genetyczny komórki musi się replikować, co związane jest z podziałem komórki na dwie potomne. Do każdej z połówek nici DNA dobudowywana jest wtedy druga nić z wykorzystaniem tej samej zasady komplementarności (uzupełniania się) zasad azotowych. Nowopowstała cząsteczka składają się więc z jednego łańcucha nowego i jednego starego. O replikacji DNA mówimy, że ma charakter semikonserwatywny (semi - łac. 'pół'). Replikacja nie jest, rzecz jasna, procesem zachodzącym spontanicznie. Wyróżniono szereg enzymów niezbędnych do jej inicjacji i dokładnego przeprowadzenia. Najważniejszy z nich to wydłużająca nową nić polimeraza DNA. Pełni ona też inną ważną rolę, kontrolując poprawność parowania zasad w powstającej nowej cząsteczce DNA. Odkrycie dokonane przez Watsona i Cricka oparte było na wcześniejszych badaniach m.in. R. Franklin, która zmarła, zanim przyznano jej nagrodę Nobla. Komplementarność zasad azotowych można wykazać, badając zawartość adeniny, tyminy, cytozyny i guaniny w próbce biologicznej. W zależności od dokładności metody analitycznej, procentowa zawartość A okaże się w przybliżeniu równa zawartości T, a C zawartości G.
Sekwencje DNA a kolejność aminokwasów w białku.
DNA przechowywany jest na ograniczonym obszarze w centrum komórki, oddzielonym od reszty cytoplazmy błoną plazmatyczną, czyli w jądrze (nie dotyczy to komórek bakterii). DNA nieustannie steruje procesami życiowymi komórki i jej reakcją na bodźce. Ekspresja (realizacja, odczytywanie) genów, a więc produkcja białek, utrzymywana jest na poziomie adekwatnym dla rodzaju komórki, dla bieżącego stanu fizjologicznego komórki, dla poziomu impulsacji z zewnątrz i zmienia się wraz z nimi. Komórka jest więc napiętym dynamicznym systemem, w którym zachodzi nieustanne odczytywanie informacji genetycznej. Pojedyncza cząsteczka DNA nie mogłaby nastarczyć z produkcją białka w wymaganym tempie. Na matrycy DNA syntetyzowane są wpierw cząsteczki kwasu rybonukleinowego. Różnią się one od DNA m.in. tym, że są jednoniciowe, jako cukier zawierają rybozę, a tyminę zastępuje podobny do niej strukturalnie uracyl (U). Cząsteczka RNA ma długość równą długości jednego genu. W szybkim tempie produkowana jest znaczna ilość cząsteczek RNA, które następnie uczestniczą w biosyntezie łańcucha białka. Stanowią wzmacniacz, który umożliwia szybkie zakumulowanie dużej ilości białkowego produktu genu.
Skąd komórka wie, jak zamienić daną sekwencję nukleotydów na łańcuch białka? Białko również jest łańcuchem, podobnie jak kwasy nukleinowe. Nie skręca się w podwójną helisę, choć niekiedy w helisę pojedynczą lub przyjmuje strukturę podobną do harmonijki. Łańcuch białka może przyjmować znacznie bardziej skomplikowane struktury, niż DNA. Można je porównać do wyplatania z włóczki figurek różnych kształtów. To, jaką rolę będzie mogło spełniać białko, czy to będzie rola szkieletu wypustki komórki nerwowej, czy struktura włosa, czy receptor w błonie czuły na sygnały hormonalne, zależy od unikalnego kształtu cząsteczki. Ogniwem w łańcuchu białka jest aminokwas. Aminokwasy w białkach połączone są specjalnym rodzajem wiązań, zwanych peptydowymi. Zmiana jednego aminokwasu w liczącej ich do kilku tysięcy cząsteczce białka przesądza o jego funkcjonalności. To, w jaki kształt zwinie się białko po syntezie wynika z sekwencji aminokwasów w jego łańcuchu. Ta kolejność jest ściśle odzwierciedlona przez kolejność nukleotydów w RNA i DNA. Oparte jest to na zasadzie, że kolejne trzy nukleotydy oznaczają jeden aminokwas białka. Kod genetyczny, czyli zasady zapisu informacji genetycznej, jest kolinearny, to znaczy kierunek zapisu informacji w kwasach nukleinowych jest zbieżny z sekwencją aminokwasów w białku. Trójki nukleotydów kodujące jeden aminokwas nazywamy tripletami albo kodonami. Ponieważ istnieją cztery rodzaje zasad azotowych, możliwe są 64 różne triplety (zasady azotowe mogą się powtarzać w triplecie). Białka buduje 20 różnych aminokwasów, więc liczba kodonów jest ponad trzykrotnie wyższa. Pewne z aminokwasów mogą być kodowane przez cztery lub nawet większą liczbą tripletów, zwykle różniących się między sobą jednym miejscem. Tę nadmiarowość nazywamy degeneracją kodu genetycznego, co w tym przypadku nie ma negatywnego wydźwięku.
Nic DNA możemy podzielić na szereg następujących po sobie kodonów, z których każdy oznacza aminokwas białka. W budowie DNA nie ma cechy, która mogłaby służyć za przecinek między kodonami. Jeżeli zdarzy się, że jeden nukleotyd w wyniku zadziałania np. któregoś z czynników mutagennych wypadnie z nici, zdarzenie to zmienia sens całej informacji genetycznej poniżej tego miejsca. Do cech kodu genetycznego zaliczamy więc również bezprzecinkowość. Znaczenie trójek nukleotydowych jest takie samo od bakterii i wirusów poczynając, a na ssakach kończąc. Oznacza to, że kod genetyczny jest w świecie żywym uniwersalny i świadczy za tezą, że powstał on w ewolucji jednorazowo. Na matrycy tego samego odcinka DNA powstanie to samo białko zasadniczo w każdej żywej komórce.
Etapy ekspresji informacji genetycznej.
Jak wspomniałem, aby informacja genetyczna mogła zostać wykorzystana, konieczne jest przepisanie jej na inny kwas nukleinowy. Nić RNA powstaje na matrycy tej z nici DNA, która nie koduje informacji i zawiera "lustrzane odbicie" sensownej sekwencji nukleotydów. Przepisywanie, czyli transkrypcję, cechuje komplementarność zasad nici matrycowej i nici syntetyzowanej, rygorystyczna kontrola poprawności syntezy dokonywana przez zespół enzymów, kierunek "czytania" sekwencji 3'→5' i kierunek "powstawania" nowej nici 5'→3'. Dowolna nić kwasu nukleinowego posiada końce 3' i 5', przy czym nici podwójnej helisy DNA skierowane są przeciwnie. Kolisty chromosom bakterii nie ma końców 3' i 5', chyba, że po przecięciu.
Drugi etap ekspresji informacji genetycznej to enzymatyczne zestawianie aminokwasów w kolejności, która odpowiada kolejności kodonów w RNA i łączenie ich ze sobą wiązaniami peptydowymi. Ten drugi etap nazywamy translacją, bo przypomina "tłumaczenie" informacji zawartej w sekwencji nukleotydów na budowę łańcucha białka. Mówi się o "dogmacie biologii molekularnej", że informacja płynie od DNA przez RNA do białka, nigdy na odwrót.
Translacja zachodzi w cytoplazmie, z tym, że u bakterii w cytoplazmie zachodzi również transkrypcja. Rozdzielenie obu kroków biosyntezy białka w przestrzeni i czasie u komórek eukariotycznych stwarza dodatkowe możliwości regulacji tempa i skali ekspresji genów. RNA jest nietrwały, toteż kopie genów nie zalegają w cytoplazmie i nie sprawiają szumu informacyjnego.
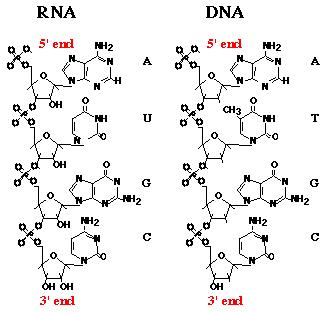
Rycina 1. Szczegóły budowy chemicznej RNA i DNA. Przedstawiono tylko połówkę podwójnej nici DNA.
Gatunki RNA.
Poza czasowym przechowywaniem informacji genetycznej, RNA spełnia w komórce liczne "techniczne" funkcje związanie z transkrypcją i translacją. Poniżej podam przykłady takich funkcji, w których specjalizują się poszczególne klasy RNA.
Matrycowy RNA (mRNA). Zwany też informacyjnym, to znany nam z wcześniejszych wywodów fragment kodujący jeden gen. mRNA przepisany z DNA zwany jest pierwotnym transkryptem. Ulega procesom dojrzewania, m.in. z sekwencji wycinane są pewne fragmenty (tzw. splicing), a oba końce transkryptu są modyfikowane chemicznie. W kompleksie z białkami mRNA przedostaje się przez pory otoczki jądrowej do cytoplazmy.
Transferowy RNA (tRNA). Krótka cząsteczka RNA o kształcie listka koniczyny lub, biorąc pod uwagę ułożenie w przestrzeni trójwymiarowej, odwróconej litery "L", ma za zadanie wychwyt aminokwasów pływających swobodnie w cytoplazmie i dostarczanie ich do tworzącego się łańcucha białka. Liczba rodzajów tRNA odpowiada liczbie kodonów. Jedna z pętli "czterolistnej koniczynki" posiada domenę (tzn. fragment cząsteczki o specyficznych właściwościach chemicznych) zwaną antykodonem, łączącą się z kodonem w mRNA. Antykodon jest więc komplementarny do kodonu.
Rybosomalny RNA (rRNA). To kwas RNA pełniący funkcje czysto strukturalne, wchodzący wraz z białkami w skład rybosomów. Są to struktury złożone z wielu białek i cząsteczek rRNA. Stanowią "platformy" łączące mRNA, enzymy i tRNA na czas syntezy łańcucha białka. Zwykle związane są z błonami plazmatycznymi, choć mogą swobodnie unosić się w cytoplazmie, jak jest np u bakterii.
Mały jądrowy RNA (snRNA). Najmniejsze z funkcjonalnych cząsteczek RNA, pełnią funkcje enzymatyczne. Uczestniczą w obróbce mRNA bezpośrednio po jego biosyntezie, a przed wysłaniem do cytoplazmy. Wycinając niepotrzebne sekwencje.
Translacja, czyli synteza polipeptydu
Aby rozpoczęła się biosynteza białka, konieczny jest adapter, który mógłby rozpoznać zarówno kodon, jak aminokwas. Jak wiemy z wcześniejszej części pracy, jest nim transportujący RNA (tRNA). Różnica właściwości chemicznych między aminokwasami a nukleotydami jest tak duża, że nie mogłyby się bezpośrednio do siebie dopasować. tRNA posiada trzy lub cztery (cecha zmienna) pętle, które zawierają zmodyfikowane, niespotykane gdzie indziej nukleotydy. Energia do tworzenia wiązań peptydowych pochodzi z samych nukleotydów, zaopatrzonych są w trzy reszty fosforanowe zamiast jednej. Pętle boczne uczestniczą w ustawieniu aminokwasu względem rybosomu, a główna pętla (antykodonowa) rozpoznaje sekwencję mRNA. Aminokwas przyłącza się do właściwego tRNA, w czym pośredniczą specyficzne enzymy należące do syntetaz aminoacylo-tRNA. Rozpoznają one specyficzny tRNA i odpowiadający mu aminokwas. Aminokwas uległ uprzednio aktywacji przez przyłączenie AMP do grupy kwasowej:
aminokwas + ATP ( aminoacylo-AMP + PP
Zaktywowany aminokwas łączy się następnie ze swoistym tRNA z udziałem enzymu:
aminoacylo-AMP + tRNA syntetaza > aminoacylo-tRNA + AMP
Godne uwagi jest, że syntetazy stanowią równie kluczowy jak tRNA element biosyntezy. Innym niezbędnym czynnikiem jest rybosom, organellum umożliwiające przestrzenne zorientowanie składników względem siebie. Rybosom składa się z podjednostki większej i mniejszej, łączących się ze sobą tylko na czas biosyntezy. Mniejsza podjednostka łączy się z transkryptem mRNA i "odnajduje" w nim kodon inicjujący biosyntezę. Jest nim zawsze kodon dla aminokwasu metioniny (AUG), jakkolwiek aminokwas ten może być w późniejszym czasie usunięty z cząsteczki, jeżeli nie pasuje do struktury danego białka. W komórkach bakterii, archeanów, mitochondriach i chloroplastach biosynteza łańcucha białka rozpoczyna się od aminokwasu formylometionina przenoszonego przez specyficzny tRNA. Następnie większa podjednostka rybosomalna przyłącza się do kompleksu i zamyka go. Do kodonu AUG zbliża się pierwszy tRNA niosący metioninę (lub formylometionimę). Gotowy rybosom posiada dwa miejsca kotwiczenia tRNA, z których jedno (miejsce A) służy jako "port" dla rozproszonych w cytoplazmie cząsteczek aminoacylo-tRNA (oczywiście tych, na które wskazuje kodon). Zsyntetyzowany dotychczas łańcuch peptydowy "przeskakuje" na nowoprzybyły aminoacylo-tRNA. Rybosom przemieszcza się o jeden kodon względem mRNA do miejsca P, a w miejscu A demonstrowany jest kolejny kodon. Mianem elongacji nazywamy wydłużanie się syntetyzowanego łańcucha polipeptydowego. Jedną cząsteczkę mRNA mogą równocześnie "odczytywać" liczne rybosomy (tworząc strukturę zwaną polisomem), co pozwala na produkcję dużej ilości produktu białkowego w jednostce czasu. Gdy w miejscu A pojawi się jeden z trzech kodonów STOP, następuje terminacja translacji. Do kodonu STOP przyłączają się zamiast tRNA specyficzne białka terminujące, które powodują oddysocjowanie łańcucha produktu polipeptydowego od rybosomu, a następnie dysocjację samego rybosomu na podjednostki. Uwolniony mRNA ulega spontanicznej degradacji.
Zsyntetyzowany na rybosomie wolny łańcuch białkowy jest następnie przekazywany do struktur wewnątrzkomórkowych, określanych mianem systemu GERL. Skrót ten jest akronimem nazw organelli: Golgi (apparathus), Endoplasmatic Reticulum, Lysosome. System kanałów, cystern i pęcherzy kieruje białko kolejno do miejsc fosforyzacji, sulfatacji bądź glikozylacji, przy czym każde białko przechodzi unikalne dla niego właśnie modyfikacje. Mogą też polegać na wycięciu fragmentów bądź skracaniu łańcucha polipeptydowego. Białko zaopatrywane jest w sekwencję adresową, decydującą o tym, dokąd zostanie przetransportowane. Część białek powstaje na potrzeby wewnętrzne komórki, część jest wydzielana w lizosomach i egzosomach.
Złożone metody badań z wykorzystaniem RTG umożliwiły zbadanie wielopoziomowej struktury białek. Najwyższym poziomem jest czwartorzędowa struktura, odnosząca się do wzajemnej konfiguracji łańcuchów polipeptydowych w białku. Role pełnione przez białka obejmują szerokie spektrum poczynając od strukturalnej (budulec włosów, rogów, kopyt, naskórka, błon komórkowych), przez szkieletową (utrzymywanie kształtu komórek, osi wypustek komórek nerwowych), wydzielniczą (hormony peptydowe, neuropeptydy, składniki śluzu), sygnalizacyjną (receptory błonowe i jądrowe, wewnątrzkomórkowe substancje transmitujące sygnał), regulacyjną (czynniki transkrypcyjne), po enzymatyczną i odpornościową. Badanie zależności między sekwencją aminokwasów w białku a ostatecznymi właściwościami cząsteczki jest przedmiotem ciągłych badań biochemików.
