Podstawową jednostką strukturalną i funkcjonalną ośrodkowego układu nerwowego jest neuron. Jest to komórka, która charakteryzuje się zdolnością do reakcji na pobudzenie. Reakcja ta polega na wytworzeniu się stanu czynnościowego, który następnie jest przekazywany dalej na inne neurony drogą synaptyczną.
Dzięki temu, że centralny system nerwowy ma zdolność odbierania i przewodzenia bodźców stąd w każdej chwili dochodzą do niego informacje na temat wszelkich zmian w zewnętrznym i wewnętrznym środowisku. Bodźce odbierane są przez receptory czuciowe, a następnie przekazywane w postaci impulsów nerwowych wędrują do centralnego systemu nerwowego gdzie są poddawane obróbce i analizie warunkując powstawanie wrażeń zmysłowych czy pobudzenie narządów wewnętrznych do pracy.
Aby zrozumieć działanie komórki nerwowej należy najpierw zapoznać się z jej budową. I tak każdy neuron składa się z ciała komórki oraz dendrytów i jednego aksonu. Przez dendryty impulsy przewodzone są w kierunku ciała komórki i aksonu. Jeśli natomiast dojdzie do sztucznego pobudzenia aksonu to impulsy mogą być przewodzone również od aksonu do ciała komórki. Akson zakończony jest rozgałęzieniem , na którego końcach znajdują się kolbki synaptyczne.
Każdy neuron otoczony jest przez błonę komórkową. Błona ta jest miejscem powstawania i przewodzenia potencjałów, których wytwarzanie jest podstawową własnością neuronów. Dzieje się tak dzięki obecności w błonie kanałów sodowych i potasowych. Kanały sodowe występują w większej ilości w okolicach wzgórka aksonu i dlatego właśnie w tym miejscu najczęściej dochodzi do powstawania potencjału czynnościowego. Tak więc kanały sodowe odpowiadają za proces depolaryzacji błony, natomiast kanały potasowe uczestniczą w procesach repolaryzacji. Pod względem fizycznym błonę komórkową można porównać do zespołu baterii sodowych i potasowych, które zostały szeregowo połączone elementami stanowiącymi opór dla przewodnictwa prądów jonowych. Opór elektryczny płynu wewnątrz i zewnątrzkomórkowego jest prawie dwa tysiące razy mniejszy od oporu samej błony komórkowej aksonu, który w spoczynku wynosi około 0.1 - 0.2 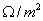 .
.
Tak duża różnica w wartości oporu jest możliwa dzięki temu, że w skład błony komórkowej wchodzą fosfolipidy będące dobrym izolatorem. Dlatego też możliwa jest izolacja dużej ilości ładunków o przeciwnym znaku przy jednoczesnym zachowaniu niewielkiej wartości potencjału spoczynkowego.
Jeżeli neuron znajduje się w stanie spoczynku to pomiędzy dwoma dowolnymi miejscami na powierzchni błony komórkowej nie występuje różnica potencjału. Mówi się, że powierzchnia tej komórki jest izopotencjalna. Jeśli natomiast dokona się pomiaru wartości różnicy potencjałów między powierzchnią komórki a jej wnętrzem to otrzyma się wartość około -60 do -90 mV. Jest to tzw. potencjał spoczynkowy. Na jego powstawanie ma wpływ właśnie różnica stężeń jonów sodowych i potasowych między środowiskiem zewnętrznym komórki a jej wnętrzem , a także przemieszczanie się tych jonów w kierunku wyznaczonym gradientem stężeń czyli dyfuzja jonów przez błonę komórkową neuronu. Błona komórkowa charakteryzuje się selektywną przepuszczalnością względem jonów sodowo - potasowych i dodatkowo jest wyposażona w pompy sodowo - potasowe. Na zewnątrz komórki stężenie jonów sodowych jest dziesięć razy większe od stężenia w środku komórki. Natomiast stężenie jonów potasowych jest trzydzieści razy większe po wewnętrznej stronie błony niż w środowisku zewnętrznym komórki.
W stanie spoczynku większość kanałów sodowych i potasowych jest zamknięta. Tak więc do dyfuzji dochodzi tylko w niewielkim stopniu i dotyczy to głównie kanałów potasowych ponieważ błona jest bardziej przepuszczalna dla tych jonów i znajduje się w niej dziesięć razy więcej kanałów dla potasu niż dla sodu.
Mimo, że tempo takiej dyfuzji jest małe to po pewnym czasie mogłoby doprowadzić do wyrównania stężeń jonów w płynie wewnątrz i zewnątrzkomórkowym i w konsekwencji spowodować zaburzenia pracy neuronu. Nad tym aby nie doszło do takiej sytuacji czuwa aktywna pompa sodowo - potasowa. Działa ona w oparciu o enzym zwany adenozynotrójfosfatazą (ATP) , który jest aktywowany właśnie przez jony sodu i potasu. Energia, która jest potrzebna do tego czynnego transportu pochodzi właśnie z hydrolizy ATP. Pompa ta transportuje jony sodu z wnętrza komórki , a jony potasu w przeciwnym kierunku. Tempo tego transportu zależy od wewnątrzkomórkowego stężenia jonów sodu i stężenia jonów potasu w płynie zewnątrzkomórkowym.
Zmiany elektryczne błony komórkowej neuronów mogą mieć charakter potencjałów lokalnych i potencjałów czynnościowych. Potencjały lokalne przemieszczają się wzdłuż błony neuronu ze spadkiem swojej amplitudy i powstają pod wpływem działania neurotransmitterów , które powodują depolaryzację lub hiperpolaryzację. Potencjały lokalne mogą ulegać sumowaniu zarówno w czasie jak i przestrzeni.
Natomiast potencjały czynnościowe pojawiają się zgodnie z zasadą " wszystko albo nic". Przemieszczają się wzdłuż błony neuronu nie zmieniając kształtu ani amplitudy. Potencjał czynnościowy powstaje w wyniku nagłej i chwilowej zmiany przepuszczalności błony i jej przewodności elektrycznej. Przewodność elektryczna błony jest odwrotnością jej oporności elektrycznej.
Przejście potencjału czynnościowego przez dany fragment neuronu powoduje powstanie chwilowego zaniku pobudliwości. Wówczas ten odcinek neuronu nie jest wrażliwy na żadne bodźce. Okres taki trwa średnio 1 milisekundę.
Czas trwania takich okresów refrakcji wskazuje ile razy w danej jednostce czasu neuron może ulec pobudzeniu . Oszacowano, że maksymalna częstotliwość pojawiania się potencjałów czynnościowych zazwyczaj nie jest większa niż 100 - 200 impulsów na jedną sekundę.
Z jednego neuronu na drugi lub z neuronu na inne komórki impulsy nerwowe są przekazywane za pośrednictwem synaps. Zakodowane w postaci impulsu informacje mogą być między synapsami przekazywane albo drogą elektryczną albo chemiczną, przy czym drugi sposób zdecydowanie dominuje.
Przekaz elektryczny polega na tym, że przemieszczający się do zakończeń aksonu potencjał czynnościowy powoduje powstanie lokalnych prądów depolaryzujących w jednym z elementów synapsy zwanym presynaptycznym. Następnie z niewielkim opóźnieniem depolaryzacja pojawia się także w elemencie postsynaptycznym. Do elektrycznego przewodnictwa synaptycznego dochodzi gdy pomiędzy neuronami istnieją kanaliki o średnicy 2 nm zwane koneksonami. Ten typ przewodzenia pojawia się stosunkowo rzadko. Jest to przewodnictwo dwukierunkowe i mało wrażliwe na działanie substancji farmakologicznych.
W większości synaps występuje więc przewodnictwo chemiczne. Związane jest ono z powstawaniem postsynaptycznego potencjału pobudzającego oraz postsynaptycznego potencjału hamującego.
Okazuje się , że można dokonywać pomiarów potencjału elektrycznego mózgu. Wykorzystuje się do tego specjalne urządzenia zwane elektroencefalografami. Pierwsze badania nad tymi urządzeniami prowadzone były w Polsce przez dwóch naukowców Adama Becka i Nikodema Cybulskiego. Niestety jak to się często zdarza pieniądze na badania skończyły się i nie mogli oni dokończyć swoich pionierskich prac. Elektroencefalograf składa się z wielu elektrod. Przeważnie ich liczba wynosi 128. Elektrody te przypinane są na głowie badanego pacjenta. Elektrody odbierają sygnały, które następnie są wzmacniane i zapisywane jako wykresy pracy mózgu.
Analiza zapisu EEG może być przydatna przy diagnozowaniu wielu chorób systemu nerwowego.
Jeśli chodzi o własności magnetyczne mózgu to ten obszar nie został jeszcze dokładnie poznany. W mózgu zlokalizowano obecność magnetytu, natomiast nie wiadomo co pełni rolę receptorów zwanych w tym przypadku magnetoreceptorami. Podejrzewa się, że magnetorecepcja odgrywała dużą rolę u naszych przodków i służył im do orientowania się w przestrzeni. Dzisiaj kiedy życie człowieka ułatwia wiele urządzeń magnetorecepcja stała się zbędna i uległa osłabieniu. Nie jest to jednak proces nieodwracalny. Prawdopodobnie jeśli człowiek zostałby skazany tylko na siebie w nieznanym środowisku zdolność magnetorecepcji ponownie uległaby zaostrzeniu.
